¹H MRS of MuscleWhat about spectroscopy of the musculoskeletal system?
|

|
Compared to its use in the brain and other body regions, ¹H-spectroscopy has had rather limited applications to date in the musculoskeletal system. As with breast lesions, high choline levels in ¹H-MRS have been used with some success to differentiate benign from malignant tumors of muscle, bones, nerves, and supporting tissues. The technique is also being increasingly used in research laboratories to study lipid deposition and fatty conversion of muscle mass in obesity and aging.
Because of shimming difficulties over large volumes, single-voxel techniques are generally preferred using a water-suppressed PRESS sequence with intermediate TE values (130−144 ms). Even in lean muscle, the ¹H spectrum is dominated by lipid resonances in the δ = 0.9−1.6 ppm range, with relatively minor peaks noted for total creatine (δ = 3.0) and choline (δ = 3.2). The lipid peaks can be further subdivided into those arising from within (intramyocellular lipids, IMCL) and between (extramyocellular lipids, EMCL) muscle cells.
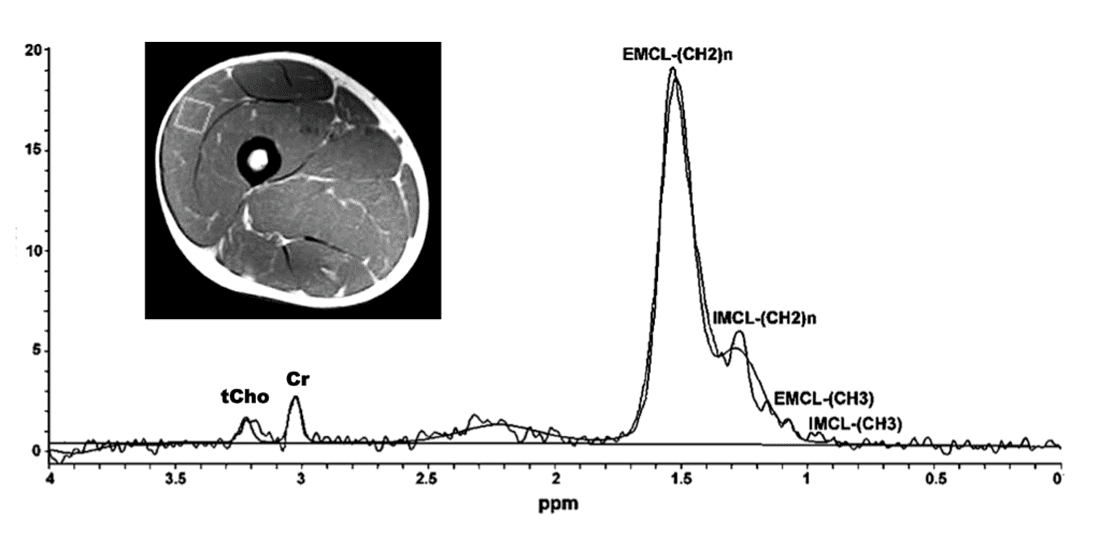
Single-voxel ¹H MRS of the vastus lateralis muscle. EMCL = Extramyocellular lipid, IMCL = Intramyocellular lipid; Cr = creatine = creatine (+ phosphocreatine), tCho = total choline = trimethylamine compounds + phosphocholine + glycerophosphocholine +
and free choline. (Adapted from Zhu et al, under CC BY license)
References
Boesch C, Slotboom J, Hoppeler H, Kreis R. In vivo determination of intra-myocellular lipids in human muscle by means of localized ¹H-MR-spectroscopy. Magn Reson Med 1997; 37:484-493.
Deshmukh S, Subhawong T, Carrino JA, Fayad L. Role of MR spectroscopy in musculoskeletal imaging. Indian J Radiol Imaging 2014; 24:210-216.
Fayad LM, Jacobs MA, Wang X, et al. Musculoskeletal tumors: How to use anatomic, functional, and
metabolic MR techniques. Radiology 2012; 265:340‑56.
Shick F, Eismann B, Jung W-I, et al. Comparison of localized proton NMR signals of skeletal muscle and fat tissue in vivo: two lipid compartments in muscle tissue. Magn Reson Med 1993; 29:158-167.
Zhu R, Wen C, Li J, et al. Lipid storage changes in human skeletal muscle during detraining. Front Physiol 2015; 6:309.
Boesch C, Slotboom J, Hoppeler H, Kreis R. In vivo determination of intra-myocellular lipids in human muscle by means of localized ¹H-MR-spectroscopy. Magn Reson Med 1997; 37:484-493.
Deshmukh S, Subhawong T, Carrino JA, Fayad L. Role of MR spectroscopy in musculoskeletal imaging. Indian J Radiol Imaging 2014; 24:210-216.
Fayad LM, Jacobs MA, Wang X, et al. Musculoskeletal tumors: How to use anatomic, functional, and
metabolic MR techniques. Radiology 2012; 265:340‑56.
Shick F, Eismann B, Jung W-I, et al. Comparison of localized proton NMR signals of skeletal muscle and fat tissue in vivo: two lipid compartments in muscle tissue. Magn Reson Med 1993; 29:158-167.
Zhu R, Wen C, Li J, et al. Lipid storage changes in human skeletal muscle during detraining. Front Physiol 2015; 6:309.
Related Questions
What peaks are seen in the ³¹P spectrum and what do they mean?
What peaks are seen in the ³¹P spectrum and what do they mean?